
Major Threats by Region
Having sketched out the main causes of species decline, we will now delve into the details. To do this, we need to drop down to the sub-national level because different parts of the country experience different patterns of anthropogenic disturbance. For our purposes, it is useful to define three broad terrestrial zones, which we will refer to throughout the text as the Agricultural South, the Industrial Forest, and the Far North (Fig. 5.5). Marine environments constitute a fourth zone. In the following sections, we will examine the major types of disturbance within each zone and describe their connection to species declines.
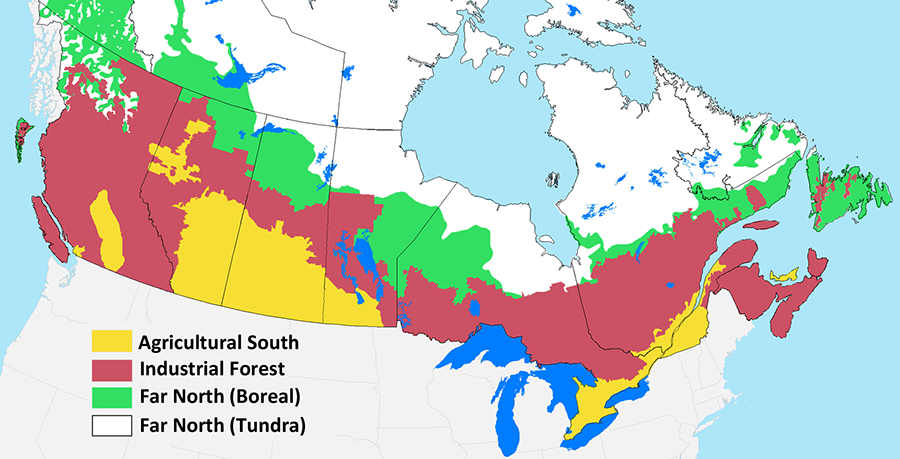
The Agricultural South
The Agricultural South zone (Fig. 5.5) includes both agricultural lands and most of Canada’s population. This region has experienced the highest rate of biodiversity decline in Canada over the past century, and the majority of Canada’s species at risk are located here (Kerr and Cihlar 2004; CESCC 2022). Not only does agriculture involve intensive land use, it has selectively targeted two specific ecosystem types: the Prairies Ecozone in the West and the Mixedwood Plains Ecozone in southern Ontario and Quebec (Fig. 2.5). No safe hinterland remains to provide habitat for the species in these two ecozones. Only 4.0% of the Prairies and 2.4% of the Mixedwood Plains exists within protected areas (ECCC 2022). Moreover, most of these protected areas are small and heavily biased to dry and unproductive rangelands (Deguise and Kerr 2006).
Habitat loss and degradation, through agricultural conversion and urban development, is the main threat to biodiversity in this region. Most of these losses occurred in the early twentieth century and, for the most part, have been permanent. Agricultural conversion is still occurring in some parts of the country, especially the interior of BC and the Peace River region of Alberta and northern BC (SC 2014b). Agricultural landscapes also feature an extremely high density of roads (Fig. 2.4). Natural disturbance regimes have been disrupted through the suppression of wildfires and the loss of native grazers (Campbell et al. 1994).

Declines in biodiversity are closely associated with the intensity of agricultural use, which has steadily increased over the years (Kerr and Cihlar 2004; Javorek and Grant 2011). As agricultural intensity increases, culminating in monoculture crop production, terrestrial ecosystems are progressively homogenized and simplified (Fig. 5.6). Wetlands in areas of intensive agriculture are also affected, through drainage, sedimentation, eutrophication, and salinization (Bartzen et al. 2010). Intensive agriculture also entails the widespread application of agricultural chemicals (Gibbs et al. 2009).
The simplified ecosystems of intensively managed agricultural lands can support only the most adaptable of species. Less adaptable species tend to concentrate in remnant patches of native habitat. Such lands account for approximately 30% of agricultural lands in the Prairie provinces, and 25% of agricultural lands in southern Ontario and Quebec (Javorek and Grant 2011). Most of the remnant patches are small and widely scattered, except in the rangelands found in the driest parts of Alberta and Saskatchewan. Many species lack the resilience needed to thrive in such fragmented and altered landscapes.
The threats to biodiversity from habitat degradation in the Agricultural South are exacerbated by agricultural chemicals and pollution, which affect both terrestrial and aquatic systems. Agricultural herbicides and pesticides can be directly toxic to wildlife species and can also reduce their food supply (Mineau et al. 2005). As a case in point, many species of native bees are currently in decline and these declines have been linked to pesticide exposure (Colla 2016). In the case of fertilizers, the problem is that a subset of species tend to benefit and become dominant, thereby decreasing overall species diversity (Haddad et al. 2000). Acid rain can cause declines in species with low acid tolerance, reducing species richness in regions with low buffering capacity (Vinebrooke et al. 2003).
Alien species present another threat to biodiversity in the Agricultural South. Most alien species are vascular plants, the bulk of which were inadvertently or deliberately introduced by immigrants from Western Europe. Today, 26% of known vascular plant species in Canada are non-native (Table 5.1). Of these, 486 are considered weedy or invasive (CFIA 2008). Many of the invasive species are agronomic grasses, such as smooth brome and crested wheatgrass, that have escaped from pastures and infiltrated native grasslands (McClay et al. 2004). Others are weeds, such as Canada thistle and leafy spurge. These alien species compete with and displace native plant species, altering community structure and function. This can have cascading effects throughout the system (Simberloff and Von Holle 1999). For example, the diversity of grassland birds is reduced where conversion to crested wheatgrass results in simpler habitat structure (Sutter and Brigham 1998). In addition, alien weeds are the cause of most herbicide use.

Invasive alien species are also a major concern in aquatic systems (Dextrase and Mandrak 2006). A wide range of taxonomic groups are involved, including plants such as purple loosestrife, invertebrates like the zebra mussel, and various non-native fish species. Invasive alien species, in combination with overfishing and pollution, are responsible for the loss of much of the original biotic community of the Great Lakes (Mandrak and Cudmore 2010). Alien species entered the Great Lakes via many routes, including the Welland Canal, the ballast water of large ships, the undersides of fishing boats, and various forms of purposeful and inadvertent release by humans (Fig. 5.7). The 185 alien species that currently reside in the Great Lakes now dominate the system (CCRM 2010).
Most other aquatic systems are not as heavily altered as the Great Lakes, but invasive species are still a concern. The zebra mussel has now invaded Lake Winnipeg, the common carp is established in several provinces, the highly invasive Prussian carp has recently been detected in southern Alberta, and purple loosestrife is widespread (Elgin et al. 2014). The deliberate and often unauthorized introduction of non-native fish species for the purpose of sport fishing has also disrupted native fish assemblages in many lakes (Chapleau et al. 1997).
Alien species also include a variety of pests and disease agents, many of which target specific hosts. For example, the fungus responsible for white-nose syndrome affects only bats. After its introduction to North America, it caused the collapse of little brown bat populations in eastern regions and is now quickly spreading westward (Frick et al. 2010). Many prominent pests, such as the emerald ash borer, gypsy moth, and the Asian long-horned beetle are invasive insects that target trees. In the future, as global warming progresses, Canada’s harsh climate will become less of a barrier for alien species, and we will see more of them enter.
Hunting, which was once a major threat to species in the agricultural zone, is much less of a concern today, though the legacy of past extirpations remains. Hunting of mammals and birds in this region is now well controlled and is mostly directed to species with large populations and high reproductive capacities, such as white-tailed deer and various waterfowl. In fact, in contrast to many other birds, waterfowl have shown a strong, increasing population trend since 1970 (NABCI 2019).
While hunting may no longer be a major concern in the Agricultural South, the same cannot be said of fishing. Recreational fishing regulations that focus on fish size and daily limits provide only a blunt instrument for managing total harvest because they do not control the total number of anglers using the resource. Fisheries involving rainbow trout in south-central BC, walleye and pike in Alberta, and lake trout in southern Ontario have all collapsed as a result of regulated recreational angling (Post et al. 2002). As a general rule, declines in recreational fish species occur in proportion to their proximity to population centres (Post et al. 2002).
The commercial harvest of freshwater fish in Canada is now considerably smaller than the recreational harvest (Post et al. 2016). However, commercial harvest was a major cause of fish declines in the past and continues to be problematic in some areas (ECO 2011). Many of the species that were once a staple, such as Atlantic salmon, lake sturgeon, and lake trout are gone from many lakes or have populations too small to support commercial harvest. As these species became depleted, the industry moved on to other species, including walleye, yellow perch, and lake whitefish. The status of these stocks is variable; some are stable, while others have collapsed (Sullivan 2003).

In addition to these widespread threats, there are threats with a regional or local impact. One of the most notable is disturbance from recreational activities, especially off-road vehicle use (Fig. 5.8). These types of disturbances increase with proximity to urban centres. They are a particular concern for at-risk plant species, the majority of which list recreation as a threat factor (McCune et al. 2013). There is also some mining and oil and gas development in the Agricultural South, but we will defer our discussion of these industries to the next section.
An important feature of all these threats is that they tend to overlap spatially. Species in the Agricultural South are rarely impacted by just one threat but by the cumulative impact of many factors operating in concert. Only the most adaptable species can thrive under such conditions.
The Industrial Forest
The Industrial Forest zone (Fig. 5.5) is defined by the spatial distribution of commercial forestry operations, which run in a broad band across the country (Fig. 2.9). Other notable forms of industrial development in this region include mining, oil and gas extraction, hydroelectric development, and peat extraction (Figs. 2.10 and 2.14).
Declines in biodiversity have been much lower in the Industrial Forest than in the Agricultural South, and there are fewer species at risk (even after accounting for lower species richness in the north; Kerr and Cihlar, 2004). It is mainly species with high sensitivity to anthropogenic disturbance that have undergone major declines, in contrast to the Agricultural South where only most adaptable species have remained stable. These sensitive species include habitat specialists, like the brown creeper (Poulin et al. 2008), and species that avoid roads and other industrial disturbances, such as woodland caribou (Dyer et al. 2002).
Species in the Industrial Forest zone have fared better than their agricultural counterparts mainly because forestry companies in most areas have long been required to reforest harvest blocks using native species. Moreover, less than half of Canada’s forests are under industrial management, leaving a large reservoir of intact forest (though most of this is in northern areas with low productivity). Finally, the pace of forestry operations is relatively slow—less than 0.5% of the total forest area is harvested each year (CFS 2022). Consequently, most forests have only been harvested once, or are still awaiting their first harvest (Venier et al. 2014).
Although habitat changes in the Industrial Forest zone are not as severe as in the Agricultural South, they are still the main cause of declines of biodiversity in this region. Forest habitat is degraded through permanent deforestation, fragmentation, changes in forest patterns and composition, and the development of roads and other infrastructure.
Deforestation currently occurs at a low rate. Since 1990, conversion to other uses has averaged approximately 50,000 ha per year, out of more than 350 million ha of total forest area (CFS 2022). However, several specific forest types have experienced large losses in the past from which they have never recovered. These include the Carolinian forests of the Great Lakes Lowlands (Suffling et al. 2003), red and white pine forests in Eastern Canada (Venier et al. 2014), deciduous forests along the northern fringe of the prairie grasslands (Timoney 2003), and Douglas fir forests in coastal BC (CCRM 2010).
The deforestation that continues to occur today is primarily from agricultural expansion and industrial development (Table 5.2). Losses to agriculture occur mainly along the northern boundary of the agricultural zone (Hobson et al. 2002). Most of the industrial impact is from oil and gas exploration and development, which is centred in northern Alberta and northeast BC (Fig. 2.10). Mining is another contributor and it is widely distributed across the country (Fig. 2.14). Sporadic, localized forest losses also occur from flooding associated with hydroelectric development. Peat mining is not included in deforestation tallies, but it constitutes another form of habitat loss in this zone (Kreutzweiser et al. 2013). Most peat mining occurs along the southern fringe of the boreal forest.
Table 5.2. The annual rate of permanent deforestation in Canada, by sector, for 2020.1
| Sector | Area (ha) |
| Agriculture | 22,378 |
| Oil and gas + mining | 15,144 |
| Infrastructure | 9,637 |
| Hydroelectric | 1,101 |
| Forestry | 1,092 |
| Total | 49,352 |
1Source: CFS 2022.
Though the rate of deforestation is low at the national scale, it can be significant at the regional scale. In some parts of northern Alberta, the annual area of forest clearing by the oil and gas sector rivals that of the forest industry (Schneider et al. 2003). Furthermore, there has been limited progress in restoring native vegetation to wells and mines after production has ceased (Osko and Glasgow 2010). Such sites have typically been restored to grass rather than forest. Consequently, forest clearing for oil and gas development and mining tends to be semi-permanent, resulting in impacts that accumulate over time (Nitschke 2008; Pickell et al. 2015).
The annual amount of permanent deforestation is dwarfed by the 700,000–800,000 ha harvested each year by the forest industry (CFS 2022). Forest harvesting is considered a transient disturbance, not deforestation, because harvest blocks must be regenerated. Nevertheless, it has widespread effects on forest structures and patterns (Grondin et al. 2018). These changes arise from the differences that exist between conventional forest harvesting and natural forms of disturbance, such as wildfire and insect attacks.
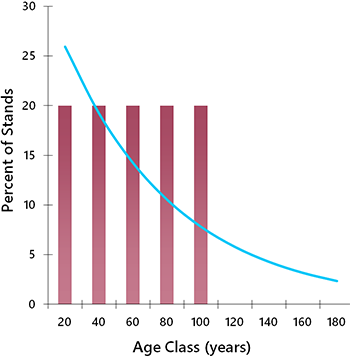
The most important difference between harvesting and natural disturbance relates to the age structure of the forest (Cyr et al. 2009). Natural disturbances are generally random, so some stands remain undisturbed for long periods simply by chance. This results in an age distribution featuring a long “tail” of older stands (Fig. 5.9). Through successional processes, the undisturbed stands develop unique structural and compositional attributes as they mature, which help to support a variety of specialist species (see Chapter 7).
In contrast, conventional forestry operations specifically target older stands because of their high wood volume and value. Moreover, fast-tracking the harvest of old stands can avert losses from fire, insect outbreak, and natural mortality, which occur at a higher rate in older age classes. Over time, the selective harvest of older stands truncates the forest age structure at the preferred harvest age (Fig. 5.9; Bergeron and Fenton 2012). This in turn causes declines in old-forest specialist species (Schmiegelow and Monkkonen 2002).
The ecological effects of age-class truncation are readily observed in forests with a long history of harvesting, such as the boreal forests of Finland. In Finland, only 6.5% of stands on managed landscapes are over 140 years old, whereas 50% of stands within protected areas are in this age group (Virkkala and Rajasarkka 2007). As might be expected, most of Finland’s threatened bird species are old-forest specialists, and they are now found mainly in protected areas. Forest age structures in Canada are not yet as skewed as those in Finland, but only because the bulk of our commercial forests were only brought into production in the last half of the twentieth century. The full story of forestry impacts in Canada has yet to be revealed.
Another type of habitat at risk from forestry is burned forest (Hannon and Drapeau 2005). In managed landscapes, salvage logging removes trees that have been burned, and fire suppression reduces the overall amount of burning. This again presents a problem for habitat specialists. In natural systems, burned stands are targeted by species like the black-backed woodpecker that exploit insects which attack dead trees.
Forest harvesting also tends to simplify forest structures and patterns (Doyon et al. 2008). Whereas fires leave standing dead trees, patches of live trees, and coarse debris, all of which contribute to habitat complexity, conventional clearcutting leaves little behind. Furthermore, in the mixed coniferous and deciduous forests of the eastern and western boreal region, silvicultural practices tend to produce pure coniferous or deciduous stands, rather than mixed stands, reducing niche complexity and species richness (Hobson and Bayne 2000; Boucher et al. 2009). Finally, conventional clearcutting results in simple spatial patterns, rather than the complex shapes and variable patch sizes produced by fire (Fig. 5.10). In all these cases, it is not the individual changes that are important, but the cumulative transformation that occurs across large areas. Simplified forests have more generalist species and fewer specialists than forests with complex niche structure (Hobson and Bayne 2000; Zhang et al. 2013).

Another threat to wildlife in the Industrial Forest zone is the vast network of roads, pipelines, and power lines that permeate the region, providing access to forestry harvest blocks, oil wells, and mines as well as routes for moving products to market (Fig. 2.4). In contrast to harvest blocks, which are reforested, roads and utility corridors are usually permanent. Moreover, new roads and corridors continue to be built each year, as remote areas are accessed for the first time.
Seismic lines, used for oil and gas exploration, add to the density of linear disturbances in forested landscapes (Fig. 5.11). In the 1980s and 1990s, an average of 46,000 km of seismic lines were approved for the forested area of Alberta each year (AEP 1998). Seismic exploration has also been intensive in northeast BC. These older lines, usually 6–8 m wide, remain as legacy disturbances today.

Exploration companies were not required to reforest these lines, and natural regeneration has been slow, often because of ongoing use by all-terrain vehicles and snowmobiles. A study of lines created in the 1960s and 1970s found that 65% remained in a cleared state 35 years after they were created, and only 8% were fully reforested (Lee and Boutin 2006). Technology now exists to create lines that are only 2.5 m wide, but it is not used consistently.
Biodiversity in the Industrial Forest is also affected by pollution, particularly from mining. Ore bodies often contain toxic elements, including cadmium, lead, and arsenic. Once the rock is ground into fine-grained tailings, these elements can migrate into the environment if not properly stored. Other types of tailings produce acid when they come in contact with oxygen and water, promoting the leaching of metals and the acidification of ground and surface waters (Shang et al. 1999). The risk of release into the environment is greatest for abandoned mines, of which there are thousands (MacKasey 2000). Accidents also happen in active mines, as demonstrated by the Mount Polley tailing pond failure in BC in 2014, which released millions of cubic metres of toxic slurry into the adjacent water system (IEEIRP 2015).
Pollution also arises from other industries. Oil and gas production causes soil and water contamination through pipeline ruptures, accidental spills, drilling discharges, and improper waste disposal. In the oil sands, toxic tailing ponds cause mortality in waterfowl and other bird species that land on them inadvertently. In the forestry sector, pulp mills release organochlorines and other processing byproducts into local waterways.
Air pollution is another common byproduct of industrial processing and can affect wide regions. One of the main threats to biodiversity is the acidification of lakes and forests through acid rain, particularly in the Shield, where little natural buffering capacity exists (Schindler 2001). The sources of SO2 and NO2 that create acid rain include coal-fired generators, industrial emissions, and vehicle emissions.
Lastly, biodiversity in the Industrial Forest is threatened by alien species. Insect pests and disease organisms, such as Dutch elm disease, white pine blister rust, and gypsy moth are of particular concern (Allen and Humble 2002). Alien plants are also a threat, especially in disturbed sites and riparian areas (Rose and Hermanutz 2004). The spread of many alien species, such as earthworms (Cameron and Bayne 2009) and agronomic grasses (Sumners and Archibold 2007), is facilitated by roads. Some of these alien species act as sporadic disturbance agents, whereas others integrate into the forest ecosystem, changing competitive relationships and altering ecological processes.
The Far North
The Far North is the largest zone, encompassing all lands north of the Industrial Forest zone (Fig. 5.5). It is not entirely pristine, but the vast majority is roadless and undisturbed (Fig. 2.4). The roads that do exist are mostly used to provide access to scattered mining developments. Some access has also been developed in association with oil and gas development, mostly in the Yukon and along the Mackenzie River in the NWT.
Since most of this region remains undisturbed, the threats to biodiversity are low relative to the southern regions. However, because of the harsh conditions in the north, resilience to disturbance is also low, and vegetation regrowth is very slow (Mallory et al. 2006). Consequently, when disturbance does occur, it has a greater effect on biodiversity than in the south.
As in the south, it is not just the footprint of individual mines and oil wells that needs to be considered, but also the cumulative effect of development at the regional scale, including the expansion of access, provision of housing for workers, and creation of other infrastructure. These cumulative effects can disrupt animal movement patterns and reduce habitat quality for species sensitive to disturbance, including caribou, grizzly bears, wolves, and wolverines (Johnson et al. 2005).
The other main threat to biodiversity in the Far North is hunting. Although Indigenous communities in the north have a long tradition of sustainable harvesting, human populations are higher today than in the past, and snowmobiles and rifles have dramatically increased the efficiency of hunting. The declines of some species, such as caribou, have been attributed to unsustainable harvest rates in combination with other factors (Boulanger et al. 2011). This is currently a point of management controversy in the Far North (Nesbitt and Adamczewski 2013; Parlee et al. 2018). Other flash points include the trophy hunting of polar bears and the traditional hunting of endangered whales (Freeman and Wenzel 2006).
Because of the remoteness and lack of access, the status of species in the Far North is difficult to determine (NABCI 2019). Most monitoring efforts have been directed to species that are socially important and relatively amenable to measurement, such as caribou and polar bears. For most other species, the impacts of human activities are not well known.
Marine
The most important threat to biodiversity in marine environments is fishing (Baum and Fuller 2016). The economic value of commercial fishing has led to conflicts between protection and exploitation, and management outcomes have generally been poor (McDevitt-Irwin et al. 2015). The collapse of the northern cod fishery is the most notorious example, but it is just one of many fisheries that have collapsed. Of 125 commercially harvested fish and invertebrate stocks with recent status assessments, only 24% are healthy (Baum and Fuller 2016). Forty marine species have been assessed by COSEWIC as endangered or threatened, but most have been denied listing under SARA on socio-economic grounds (McDevitt-Irwin et al. 2015).
Fishing has several distinct effects on marine species. Direct mortality from harvesting is the most obvious and can result in the collapse of targeted species if the rate of harvest is too high. The collapse of targeted fish stocks can in turn affect other species by disrupting the normal food chain (Fauchald 2010). Commercial fishing also causes collateral damage through the inadvertent harvest of non-target species (bycatch). In the Pacific groundfish trawl fishery, discarded bycatch amounted to 20% of the total biomass caught between 1996 and 2006 (Driscoll et al. 2009). Finally, fishing methods based on bottom trawling can destroy seafloor habitats. This has been identified as a major threat to cold water corals and sponges, which are found off Canada’s eastern and western coasts (Wallace et al. 2015).

Another threat to species in marine environments is pollution. Most industrial and residential liquid waste eventually makes its way into rivers that discharge into the ocean. Thus, oceans become contaminated with myriad chemical products. These chemicals are rapidly diluted, to be sure. Even so, some enter the food chain and undergo bioamplification. In higher-level predators, like killer whales, exposure can reach toxic levels (Alava et al. 2016). Ocean contamination with plastics is also a growing concern, though the population-level effects are not yet understood (Rochman et al. 2016). Finally, catastrophic pollution occasionally happens through oil spills from tankers and offshore drilling installations (Fig. 5.12). This has been one of the main points of resistance to the construction of the Trans Mountain pipeline from Alberta to the BC coast.
For some species, particularly whales, collisions with ships and entanglement in fishing gear are significant threats (Williams and O’Hara 2010). Incidence is highest along the southern coast of BC and in the Gulf of St. Lawrence, where whales must contend with high levels of maritime traffic. For the North Atlantic right whales, the issue has become critical, with 12 whales killed in the Gulf of St. Lawrence 2017. Most died from blunt trauma from ship strikes, though at least two died from entanglement in fishing gear (Daoust et al. 2018).
