
Hearing and the Other Senses
Learning Objectives
- Describe the basic anatomy and function of the auditory system
- Explain how we encode and perceive pitch
- Discuss how we localize sound
- Describe the basic functions of the chemical senses
- Explain the basic functions of the somatosensory, nociceptive, and thermoceptive sensory systems
- Describe the basic functions of the vestibular, proprioceptive, and kinesthetic sensory systems
Our auditory system converts pressure waves into meaningful sounds. This translates into our ability to hear the sounds of nature, to appreciate the beauty of music, and to communicate with one another through spoken language. This section will provide an overview of the basic anatomy and function of the auditory system. It will include a discussion of how the sensory stimulus is translated into neural impulses, where in the brain that information is processed, how we perceive pitch, and how we know where sound is coming from.
Anatomy of the Auditory System
The ear can be separated into multiple sections. The outer ear includes the pinna, which is the visible part of the ear that protrudes from our heads, the auditory canal, and the tympanic membrane, or eardrum. The middle ear contains three tiny bones known as the ossicles, which are named the malleus (or hammer), incus (or anvil), and the stapes (or stirrup). The inner ear contains the semi-circular canals, which are involved in balance and movement (the vestibular sense), and the cochlea. The cochlea is a fluid-filled, snail-shaped structure that contains the sensory receptor cells (hair cells) of the auditory system (Figure 5.25).
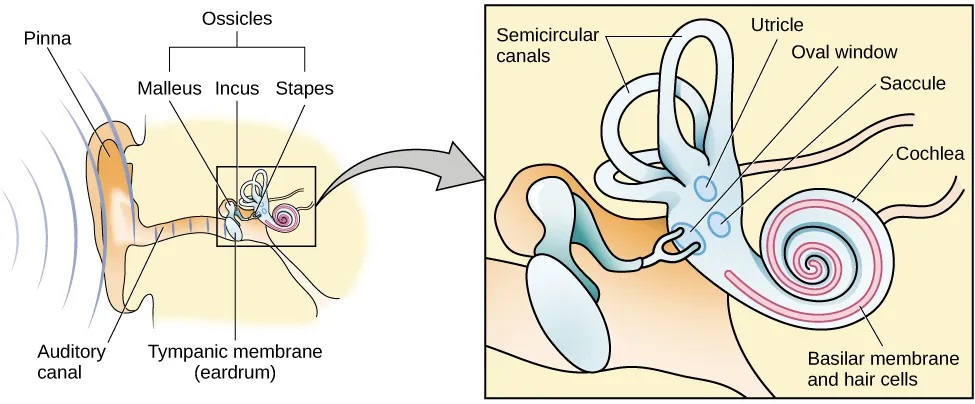
Sound waves travel along the auditory canal and strike the tympanic membrane, causing it to vibrate. This vibration results in movement of the three ossicles. As the ossicles move, the stapes presses into a thin membrane of the cochlea known as the oval window. As the stapes presses into the oval window, the fluid inside the cochlea begins to move, which in turn stimulates hair cells, which are auditory receptor cells of the inner ear embedded in the basilar membrane. The basilar membrane is a thin strip of tissue within the cochlea.
The activation of hair cells is a mechanical process: the stimulation of the hair cell ultimately leads to activation of the cell. As hair cells become activated, they generate neural impulses that travel along the auditory nerve to the brain. Auditory information is shuttled to the inferior colliculus, the medial geniculate nucleus of the thalamus, and finally to the auditory cortex in the temporal lobe of the brain for processing. Like the visual system, there is also evidence suggesting that information about auditory recognition and localization is processed in parallel streams (Rauschecker & Tian, 2000; Renier et al., 2009).
Pitch Perception
Different frequencies of sound waves are associated with differences in our perception of the pitch of those sounds. Low-frequency sounds are lower pitched, and high-frequency sounds are higher pitched. How does the auditory system differentiate among various pitches?
Several theories have been proposed to account for pitch perception. We’ll discuss two of them here: temporal theory and place theory. The temporal theory of pitch perception asserts that frequency is coded by the activity level of a sensory neuron. This would mean that a given hair cell would fire action potentials related to the frequency of the sound wave. While this is a very intuitive explanation, we detect such a broad range of frequencies (20–20,000 Hz) that the frequency of action potentials fired by hair cells cannot account for the entire range. Because of properties related to sodium channels on the neuronal membrane that are involved in action potentials, there is a point at which a cell cannot fire any faster (Shamma, 2001).
The place theory of pitch perception suggests that different portions of the basilar membrane are sensitive to sounds of different frequencies. More specifically, the base of the basilar membrane responds best to high frequencies and the tip of the basilar membrane responds best to low frequencies. Therefore, hair cells that are in the base portion would be labeled as high-pitch receptors, while those in the tip of basilar membrane would be labeled as low-pitch receptors (Shamma, 2001).
In reality, both theories explain different aspects of pitch perception. At frequencies up to about 4000 Hz, it is clear that both the rate of action potentials and place contribute to our perception of pitch. However, much higher frequency sounds can only be encoded using place cues (Shamma, 2001).
Sound Localization
The ability to locate sound in our environments is an important part of hearing. Localizing sound could be considered similar to the way that we perceive depth in our visual fields. Like the monocular and binocular cues that provided information about depth, the auditory system uses both monaural (one-eared) and binaural (two-eared) cues to localize sound.
Each pinna interacts with incoming sound waves differently, depending on the sound’s source relative to our bodies. This interaction provides a monaural cue that is helpful in locating sounds that occur above or below and in front or behind us. The sound waves received by your two ears from sounds that come from directly above, below, in front, or behind you would be identical; therefore, monaural cues are essential (Grothe, Pecka, & McAlpine, 2010).
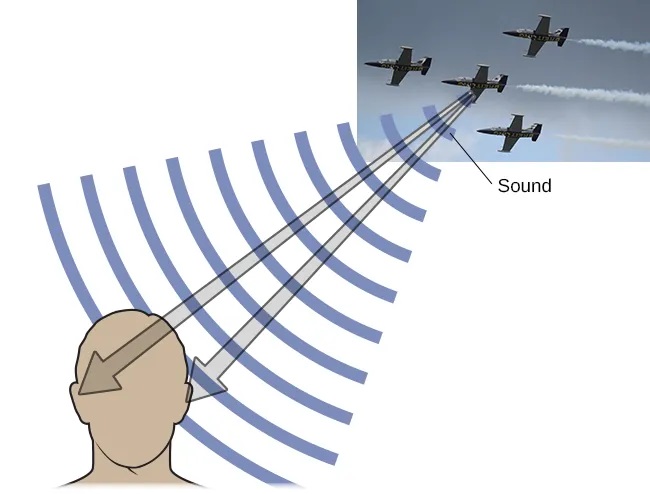
Binaural cues, on the other hand, provide information on the location of a sound along a horizontal axis by relying on differences in patterns of vibration of the eardrum between our two ears. If a sound comes from an off-center location, it creates two types of binaural cues: interaural level differences and interaural timing differences. Interaural level difference refers to the fact that a sound coming from the right side of your body is more intense at your right ear than at your left ear because of the attenuation of the sound wave as it passes through your head. Interaural timing difference refers to the small difference in the time at which a given sound wave arrives at each ear (Figure 5.26). Certain brain areas monitor these differences to construct where along a horizontal axis a sound originates (Grothe et al., 2010).
Hearing Loss
Deafness is the partial or complete inability to hear. Some people are born without hearing, which is known as congenital deafness. Other people suffer from conductive hearing loss, which is due to a problem delivering sound energy to the cochlea. Causes for conductive hearing loss include blockage of the ear canal, a hole in the tympanic membrane, problems with the ossicles, or fluid in the space between the eardrum and cochlea. Another group of people suffer from sensorineural hearing loss, which is the most common form of hearing loss. Sensorineural hearing loss can be caused by many factors, such as aging, head or acoustic trauma, infections and diseases (such as measles or mumps), medications, environmental effects such as noise exposure (noise-induced hearing loss, as shown in Figure 5.27), tumors, and toxins (such as those found in certain solvents and metals).

Given the mechanical nature by which the sound wave stimulus is transmitted from the eardrum through the ossicles to the oval window of the cochlea, some degree of hearing loss is inevitable. With conductive hearing loss, hearing problems are associated with a failure in the vibration of the eardrum and/or movement of the ossicles. These problems are often dealt with through devices like hearing aids that amplify incoming sound waves to make vibration of the eardrum and movement of the ossicles more likely to occur.
When the hearing problem is associated with a failure to transmit neural signals from the cochlea to the brain, it is called sensorineural hearing loss. One disease that results in sensorineural hearing loss is Ménière’s disease. Although not well understood, Ménière’s disease results in a degeneration of inner ear structures that can lead to hearing loss, tinnitus (constant ringing or buzzing), vertigo (a sense of spinning), and an increase in pressure within the inner ear (Semaan & Megerian, 2011). This kind of loss cannot be treated with hearing aids, but some individuals might be candidates for a cochlear implant as a treatment option. Cochlear implants are electronic devices that consist of a microphone, a speech processor, and an electrode array. The device receives incoming sound information and directly stimulates the auditory nerve to transmit information to the brain.
WHAT DO YOU THINK? Deaf Culture
In the United States and other places around the world, deaf people have their own language, schools, and customs. This is called deaf culture. In the United States, deaf individuals often communicate using American Sign Language (ASL); ASL has no verbal component and is based entirely on visual signs and gestures. The primary mode of communication is signing. One of the values of deaf culture is to continue traditions like using sign language rather than teaching deaf children to try to speak, read lips, or have cochlear implant surgery.
When a child is diagnosed as deaf, parents have difficult decisions to make. Should the child be enrolled in mainstream schools and taught to verbalize and read lips? Or should the child be sent to a school for deaf children to learn ASL and have significant exposure to deaf culture? Do you think there might be differences in the way that parents approach these decisions depending on whether or not they are also deaf?
Vision and hearing have received an incredible amount of attention from researchers over the years. While there is still much to be learned about how these sensory systems work, we have a much better understanding of them than of our other sensory modalities. In this section, we will explore our chemical senses (taste and smell) and our body senses (touch, temperature, pain, balance, and body position).
The Chemical Senses
Taste (gustation) and smell (olfaction) are called chemical senses because both have sensory receptors that respond to molecules in the food we eat or in the air we breathe. There is a pronounced interaction between our chemical senses. For example, when we describe the flavor of a given food, we are really referring to both gustatory and olfactory properties of the food working in combination.
Taste (Gustation)
You have learned since elementary school that there are four basic groupings of taste: sweet, salty, sour, and bitter. Research demonstrates, however, that we have at least six taste groupings. Umami is our fifth taste. Umami is actually a Japanese word that roughly translates to yummy, and it is associated with a taste for monosodium glutamate (Kinnamon & Vandenbeuch, 2009). There is also a growing body of experimental evidence suggesting that we possess a taste for the fatty content of a given food (Mizushige, Inoue, & Fushiki, 2007).
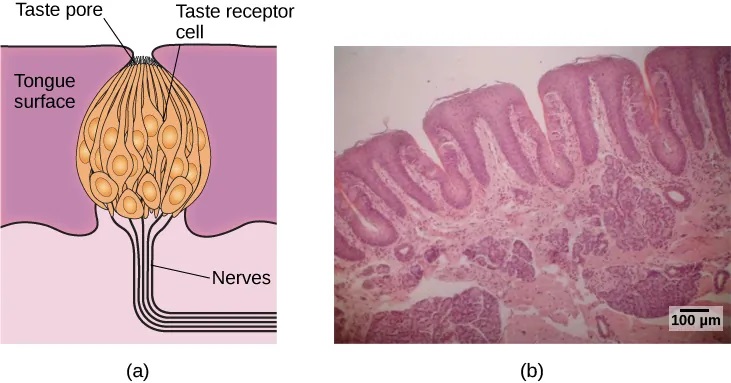
Molecules from the food and beverages we consume dissolve in our saliva and interact with taste receptors on our tongue and in our mouth and throat. Taste buds are formed by groupings of taste receptor cells with hair-like extensions that protrude into the central pore of the taste bud (Figure 5.28). Taste buds have a life cycle of ten days to two weeks, so even destroying some by burning your tongue won’t have any long-term effect; they just grow right back. Taste molecules bind to receptors on this extension and cause chemical changes within the sensory cell that result in neural impulses being transmitted to the brain via different nerves, depending on where the receptor is located. Taste information is transmitted to the medulla, thalamus, and limbic system, and to the gustatory cortex, which is tucked underneath the overlap between the frontal and temporal lobes (Maffei, Haley, & Fontanini, 2012; Roper, 2013).
Smell (Olfaction)
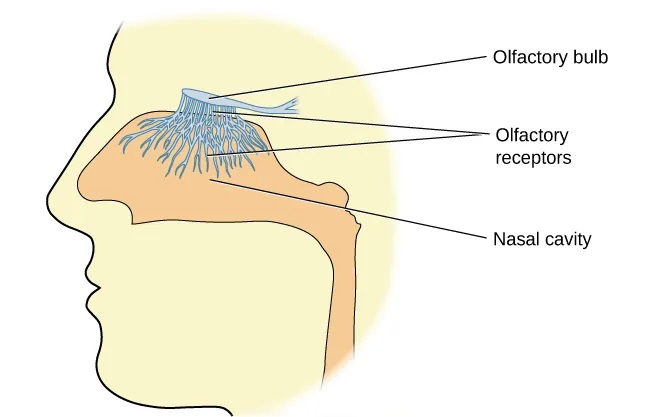
Olfactory receptor cells are located in a mucous membrane at the top of the nose. Small hair-like extensions from these receptors serve as the sites for odor molecules dissolved in the mucus to interact with chemical receptors located on these extensions (Figure 5.29). Once an odor molecule has bound a given receptor, chemical changes within the cell result in signals being sent to the olfactory bulb: a bulb-like structure at the tip of the frontal lobe where the olfactory nerves begin. From the olfactory bulb, information is sent to regions of the limbic system and to the primary olfactory cortex, which is located very near the gustatory cortex (Lodovichi & Belluscio, 2012; Spors et al., 2013).
There is tremendous variation in the sensitivity of the olfactory systems of different species. We often think of dogs as having far superior olfactory systems than our own, and indeed, dogs can do some remarkable things with their noses. There is some evidence to suggest that dogs can “smell” dangerous drops in blood glucose levels as well as cancerous tumors (Wells, 2010). Dogs’ extraordinary olfactory abilities may be due to the increased number of functional genes for olfactory receptors (between 800 and 1200), compared to the fewer than 400 observed in humans and other primates (Niimura & Nei, 2007).
Many species respond to chemical messages, known as pheromones, sent by another individual (Wysocki & Preti, 2004). Pheromonal communication often involves providing information about the reproductive status of a potential mate. So, for example, when a female rat is ready to mate, she secretes pheromonal signals that draw attention from nearby male rats. Pheromonal activation is actually an important component in eliciting sexual behaviour in the male rat (Furlow, 1996, 2012; Purvis & Haynes, 1972; Sachs, 1997). There has also been a good deal of research (and controversy) about pheromones in humans (Comfort, 1971; Russell, 1976; Wolfgang-Kimball, 1992; Weller, 1998).
Touch, Thermoception, and Nociception
A number of receptors are distributed throughout the skin to respond to various touch-related stimuli (Figure 5.30). These receptors include Meissner’s corpuscles, Pacinian corpuscles, Merkel’s disks, and Ruffini corpuscles. Meissner’s corpuscles respond to pressure and lower frequency vibrations, and Pacinian corpuscles detect transient pressure and higher frequency vibrations. Merkel’s disks respond to light pressure, while Ruffini corpuscles detect stretch (Abraira & Ginty, 2013).
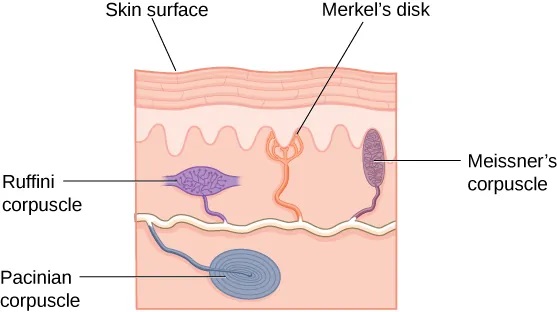
In addition to the receptors located in the skin, there are also a number of free nerve endings that serve sensory functions. These nerve endings respond to a variety of different types of touch-related stimuli and serve as sensory receptors for both thermoception (temperature perception) and nociception (a signal indicating potential harm and maybe pain) (Garland, 2012; Petho & Reeh, 2012; Spray, 1986). Sensory information collected from the receptors and free nerve endings travels up the spinal cord and is transmitted to regions of the medulla, thalamus, and ultimately to somatosensory cortex, which is located in the postcentral gyrus of the parietal lobe.
Pain Perception
Pain is an unpleasant experience that involves both physical and psychological components. Feeling pain is quite adaptive because it makes us aware of an injury, and it motivates us to remove ourselves from the cause of that injury. In addition, pain also makes us less likely to suffer additional injury because we will be gentler with our injured body parts.
Generally speaking, pain can be considered to be neuropathic or inflammatory in nature. Pain that signals some type of tissue damage is known as inflammatory pain. In some situations, pain results from damage to neurons of either the peripheral or central nervous system. As a result, pain signals that are sent to the brain get exaggerated. This type of pain is known as neuropathic pain. Multiple treatment options for pain relief range from relaxation therapy to the use of analgesic medications to deep brain stimulation. The most effective treatment option for a given individual will depend on a number of considerations, including the severity and persistence of the pain and any medical/psychological conditions.
Some individuals are born without the ability to feel pain. This very rare genetic disorder is known as congenital insensitivity to pain (or congenital analgesia). While those with congenital analgesia can detect differences in temperature and pressure, they cannot experience pain. As a result, they often suffer significant injuries. Young children have serious mouth and tongue injuries because they have bitten themselves repeatedly. Not surprisingly, individuals suffering from this disorder have much shorter life expectancies due to their injuries and secondary infections of injured sites (U.S. National Library of Medicine, 2013).
The Vestibular Sense, Proprioception, and Kinesthesia
The vestibular sense contributes to our ability to maintain balance and body posture. As Figure 5.31 shows, the major sensory organs (utricle, saccule, and the three semicircular canals) of this system are located next to the cochlea in the inner ear. The vestibular organs are fluid-filled and have hair cells, similar to the ones found in the auditory system, which respond to movement of the head and gravitational forces. When these hair cells are stimulated, they send signals to the brain via the vestibular nerve. Although we may not be consciously aware of our vestibular system’s sensory information under normal circumstances, its importance is apparent when we experience motion sickness and/or dizziness related to infections of the inner ear (Khan & Chang, 2013).
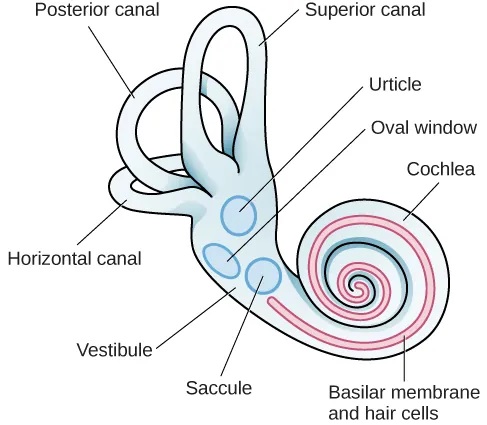
In addition to maintaining balance, the vestibular system collects information critical for controlling movement and the reflexes that move various parts of our bodies to compensate for changes in body position. Therefore, both proprioception (perception of body position) and kinesthesia (perception of the body’s movement through space) interact with information provided by the vestibular system.
These sensory systems also gather information from receptors that respond to stretch and tension in muscles, joints, skin, and tendons (Lackner & DiZio, 2005; Proske, 2006; Proske & Gandevia, 2012). Proprioceptive and kinesthetic information travels to the brain via the spinal column. Several cortical regions in addition to the cerebellum receive information from and send information to the sensory organs of the proprioceptive and kinesthetic systems.
